

Eskandari N, Azarbayjani M A, Matinhomaee H, Fatolahi H. The Effect of Date Pollen Extract/Testosterone and Resistance Training on Gene Expression and Synthesis Rate of Occludin and Cadherin Proteins of Blood-Spinal Cord Barrier in Male Rats. cmja 2023; 13 (3) :11-20
URL:
http://cmja.arakmu.ac.ir/article-1-946-fa.html
اسکندری گودرزی نیلوفر، آذربایجانی محمدعلی، متین همایی حسن، فتح اللهی حسین. اثر عصاره گرده خرما/تستوسترون و تمرین مقاومتی بر بیان ژن و میزان سنتز پروتئینهای اوکلودین و کادهرین سد خونی- نخاعی رتهای نر. فصلنامه طب مکمل. 1402; 13 (3) :11-20
URL: http://cmja.arakmu.ac.ir/article-1-946-fa.html
1- گروه فیزیولوژی ورزشی، واحد تهران مرکزی، دانشگاه آزاد اسلامی، تهران، ایران
2- گروه فیزیولوژی ورزشی، واحد تهران مرکزی، دانشگاه آزاد اسلامی، تهران، ایران ، m_azarbayjani@iauctb.ac.ir
3- گروه تربیت بدنی، واحد پردیس، دانشگاه آزاد اسلامی، پردیس، ایران
متن کامل [PDF 1532 kb]
(952 دریافت)
|
چکیده (HTML) (2080 مشاهده)
متن کامل: (2383 مشاهده)
مقدمه
در انسان و پستانداران ارتباط کنترل شده رگهای خونی با اجزای سیستم عصبی مرکزی از الگوی تقریباً یکسانی پیروی مینمایید. در این ارتباط فیزیولوژیکی، سه رابط تخصصی امکان ورود انتخابی مواد مغذی، ویتامینها، یونها، لیپیدها و مولکولهای کوچک را از جریان خون به مغز (سد خونی- مغزی؛BBB)، مایع مغزی نخاعی (BCSFB) و سد خونی- طناب نخاعی (BSCB) ممکن میسازند (1). این سدهای فیزیکی نقش مهمی در حفاظت سیستم عصبی در برابر عوامل خارجی مانند هورمونها، سموم و پاتوژن ها دارند. بنابراین، هرگونه تغییر ناشی از جهش یا مکانیسم سیگنال دهی مستقیماً منجر به جابجایی پاسخهای التهابی- عصبی و تغییر انسجام این سدها میشود (2).
با اینکه ترتیبات آناتومیکی سد خونی- نخاعی مشابه با سد خونی- مغزی است؛ وجود چند اختلاف ساختاری مانند تعداد کم پری سیت ها، ترکیب پروتئینی مویرگها همراه با نفوذپذیری بیشتر سیتوکین هایی مانند IFN-α/γ و TNF-αاز دلایل نفوذپذیری و جذب بیشتر بافت نخاع نسبت به مغز محسوب میشود (3). بر این اساس، پاسخهای ایمنی نخاع هنگام اختلال از نظر بالینی متفاوت از پاسخهای مغزی خواهد بود. این پدیده در بیماریهای مختلف مرتبط با نخاع مانند اختلال تهاجم پری عصبی، اختلال عملکرد مفصل ساکروایلیاک و آسیب طناب نخاعی دیده میشود (4). وجود این تفاوتها ضرورت مطالعه و بررسی بیشتر سد خونی- نخاعی را در تحقیقات بالینی نشان میدهد.
سد پویای BSCB توسط بافت های مختلف و بسیاری از ماتریسهای خارج سلولی حمایت میشود. در این مجموعه منظم، عمکرد توسط ترکیبی از ساختارهای پروتئینی شامل اتصالات محکم (TJs) و اتصالات چسبنده (AJs) تنظیم میشود. پروتئینهای موجود در سیستم TJ که عمدتاً شامل اوکلودین و کلودین 1، 2، 5، 8، 13 میشوند، در تنظیم ارتباطات بین سلولی و حمل و نقل پاراسلولی نقش اساسی دارند (5). اوکلودین به عنوان اولین پروتیین غشایی انتگرال مجموعه یکپارچه سلولهای اپیتلیال و اندوتلیال اهمیت فراوانی در ایجاد متاستاز سلولهای سرطانی دارد. با اینکه مکانیسم عمل این پروتئین به روشنی مشخص نیست، ظاهراً طیف وسیعی از فرآیندهای سیگنالینگ از جمله فعالسازی پروتئین کیناز سی، GTPases و فعال شدن سایتوکاین ها در تنظیم آن نقش دارند (6). با اینحال اهمیت اوکلودین در کاهش نفوذپذیری سد خونی- نخاعی در مدلهای مختلف آزمودنی براساس میزان التهاب مزمن و عدم یکپارچگی آن نشان داده شده است (7). از طرفی مولکول چسبنده یا اتصالی کادهرین با اتصال به اسکلت سلولی علاوه بر تشکیل بافت های منجمد نقش مهمی در ایجاد پشتیبانی و تمایز عملکرد گیرندههای عصبی و مسیرهای سیگنالی تنظیم ژن دارد. این پروتئین که برای عملکرد خود به یون کلسیم (Ca2+) وابسته است، با تنظیم سطح تماس و پایداری نقش مهمی در مورفوژنز و هوموستاز بافت دارد. این شرایط نمایانگر این است که تغییر خصوصیات کادهرین ها مستقیماً با طیف گستردهای از بیماریهای خود ایمن و مجامع چند سلولی مرتبط است (4, 8). بنابراین تجزیه و تحلیل دقیق الگوهای بیان این پروتئین اجازه بررسی فرضیههای مربوط به بهبود عملکرد CNS را خواهد داد.
اگرچه بررسی تغییرات سازهای سد BBB تحت تأثیر اختلالات مولتیپل اسکلروزیس (MS)، بیماری آلزایمر و پارکینسون بینشهای ارزندهای در مورد چگونگی یکپارچگی و عملکرد این اتصالات ارائه داده است، تحقیقات عملکردی مربوط به نقش سد BSCB در آسیبهای تروماتیک و غیر تروماتیک مرتبط با افزایش سن و سرطان محدود است (9). از طرفی راهکارهای تقویت سازی این ساختارها در مدلهای انسانی و حیوانی سالم با محدودیتهای پژوهشی زیادی همراه است. در واقع نقش مداخلات به عنوان مهمترین عامل محیطی غیرتهاجمی پیشگیری کننده بر مدولاسیون مسیرهای خاص یا اجزای مولکولی/ پروتئینی این سد مشخص نیست (9, 10). در این راستا درخت نخل با محصولاتی مانند گرده خرمـا، میوه و سلولهای بنیـادی سرشار از انواع ترکیبات مؤثر مانند آلکالوئیدها، تانن ها، فلاونوئیدها، ترپن هـا و قنـد است. عصاره گرده خرما (DPP) مخلوط گیاهی سلولهای زایای نر گلهای نخل است که به طور سنتی به عنوان یک مکمل غذایی تقویت کننده قدرت باروری دو جنس مورد استفاده قرار میگیرد (11). گزارش شده است که دانههای این گرده حاوی مقادیر قابل توجهی ویتامین، انواع مواد معدنی و اسیدهای آمینه به علاوه ترکیبات استروژنن، کلسترول و استرادیول است (12). اگرچه اسـتفاده از مکمـل ایـن گیــاه قادر به بهبود شاخصهای التهــابی ناشی از کــوفتگی عضلانی نبوده است (13)، عصاره آبی آن با متعادل سازی سطوح هورمونی و گلوکز، خنثی کردن سموم و بهبود عملکرد کبد قادر به کاهش میزان فشار اکسایشی است (14). بنابراین از نقطه نظر تئوری منطقی به نظر میرسد میزان کارایی این عصاره در ارتقای عملکرد پروتئینهای بافت نخاع مورد بررسی قرار گیرد.
در کنار مکمل گرده خرما، افزایش سطح فعالیت بدنی یکی از عوامل مطرح در هوموستاز هورمونها، میزان تراکم بافت چربی، بهبود عملکرد قلب و کبد، کنترل سطح کلسترول و گلوکز خون است. در واقع فعالیت بدنی منظم یک استراتژی غیردارویی امیدوارکننده برای تعدیل ردوکس، بهبود متابولیسم و افزایش ترشح آدیپوکین محسوب میشود (15). از این منظر، یکی از جنبههای مبهم کنونی ارتباط فعالیتهای بدنی براساس متغیرهای شکل و مدت تمرین با میزان نفوذپذیری BSCB است (9). در این راستا تمرینات مقاومتی شامل تمرینات با وزنه و تمرینات توانی موجب سازگاریهای فیزیولوژیکی متفاوتی میشود. این تمرینات با سرکوب فاکتورهای التهابی TNF α، IL 6، IL1 β، و MCP 1 و افزایش آنزیمهای آنتی اکسیداتیو یک اثر پیشگیرانه دارند (13, 16). اینکه تمرینات مقاومتی بواسطه مکانیسمهای ذکر شده همراه با کاهش گلوتاتیون سبب چه تغییراتی در پروتین هــای اتصال محکم و اتصال چسبنده سد خونی- نخاعی میشود، نیاز به بررسی دارد. از طرفی، مصرف مکملهای آنابولیک- آندروژنی (AAS) عموماً هنگام تمرینات مقاومتی مجاز است. چنین تصور میشود که تستوسترون از طریق الزام گیرندههای خاص آندروژنی بر عروق نخاعی اثرگذار باشد (17). با اینحال میزان اثرگذاری آن بر سایر پروتئینهای سد نخاعی- خونی مشخص نیست. لذا لزوم بررسی اثرات همزمان مصرف تستوسترون بر تغییرات ژنتیکی و ظرفیت سنتز مولکولی این سد قابل توجیه است.
در مجموع، با توجه به فقدان دادههای مربوط به اثر تستوسترون/ مکمل گرده خرما و تمرین مقاومتی بر میزان تغییرات پروتئینهای TJ و AJ سد نخاعی- خونی، لزوم شناسایی میزان اثر یا تداخل این مداخلات ضروری به نظر میرسد. در این رابطه ارزیابی موردی میزان بیان ژن یا میزان سنتز پروتئینهای ویژه نخاعی یکی از محدودیتهای پژوهشی حوزه تجزیه و تحلیل سلولی تلقی میشود (18). بر این اساس در این مطالعه اثر چهار هفته مصرف عصاره گرده خرما، تستوسترون و تمرین مقاومتی بر میزان بیان ژن و بیان پروتئینهای اوکلودین و کادهرین بافت نخاع مورد ارزیابی قرار گرفته است. نتایج این مطالعه اطلاعات با ارزشی در رابطه با میزان نفوذپذیری سد BSCB در اختیار متخصصان علوم ورزشی، پزشکی، تغذیه و سایر گروههای حوزه درمان آسیبهای نخاعی قرار خواهد داد.
روش کار
در یک مگاپروژه تجربی با طرح پس آزمون/گروه کنترل تعداد 30 رت نر نژاد ویستار با دامنه سنی 8 تا 10 هفته و وزن 220- 200 گرم از انستیتو پاستور تهران تهیه گردید. این آزمودنیها پس از انتقال به پانسیون آزمایشگاه هیستوژنوتک در قفسهایی با سایز استاندارد و چرخه روشنایی 12 ساعته با دسترسی آزاد به آب و غذای ویژه در 6 گروه کنترل؛ تمرین مقاومتی؛ عصاره گرده خرما (DPP)؛ تستوسترون سـنتتیک؛ عصاره DPP و تمرین مقاومتی؛ تستوسترون و تمرین مقاومتی در محیطی با دمای 3 ±22 درجه سانتیگراد نگهداری شدند. کلیه مراحل آزمایشگاهی براساس دستورالعمل تشکیل، سطحبندی و شرح وظایف کمیتههای اخلاق در پژوهشهای زیستپزشکی با کد IR.UT.SPORT.REC.1397.028 انجام شد.
در گروه تمرین مقاومتی چهار هفته تمرین فزاینده بر روی نردبان 110 سانتی متری دو کاناله با مشخصات پنجاه پله 50 سانتی متری با فواصل 2 سانتی متری و شیب 80 درجه انجام گردید. در این پروتکل از وزنههای سربی ماهی گیری چسبیده شده به دم رت ها (2-1 سانتی متر بعد از محل رویش مو)، به عنوان مقاومت استفاده شد. در این راستا پس از دو هفته آموزش و آشناسازی، تمرین در هفته اول با وزنه 40 درصد وزن حیوانات شروع و سپس تا 75 درصد وزن آنها در پایان هفته چهارم ادامه یافت. بدین منظور رت ها ابتدا در پایین پله قرار گرفته و سپس توسط ضربات بسیار آهسته یا لمس دم آنها جهت بالا رفتن برانگیخته میشدند. از اینرو، در این مطالعه از هیچگونه پاداش و تحریک غیرطبیعی مانند تحریک الکتریکی، آب سرد یا فشار هوا استفاده نشد. یک تکرار موفق زمانی بود که حیوان بتواند در زمان 8 ثانیه به صورت کامل تمام پلهها را بالا برود. تعداد تکرارها در هر جلسه 8 تا 12 تکرار در نظر گرفته شد. وقتی آزمودنی به بالای نردبان میرسید بعد از 2 دقیقه استراحت برای تکرار بعدی آماده میشد. به منظور گرم کردن و سرد کردن حیوانات از فرمول 5 بار بالا رفتن بدون وزنه از نردبان استفاده شد (13, 19).
جهت تهیه عصاره DPP مقادیر کافی از پودر گرده نخلهای نر شهر شهداد استان کرمان تهیه و تا زمان استفاده در یخچالهای پژوهشکده گیاهان دارویی جهاد دانشگاهی استان البرز نگهداری شد. در فرآیند عصاره گیری ابتدا 250 گرم از گرده مورد نظر در دستگاه پرکولاتور (مدل 10lit، شرکت فناوران بایامد) ریخته شد. سپس در سه بار تکرار محلولی به میزان 670 میلی لیتر در ترکیب با اتانول 90 درصد بدست آمد. این عصارهها توسط دستگاه تقطیر در خلأ تغلیظ و حلال آن بطور کامل حذف گردید. میزان بازده عصاره گیری 25/85 درصد بدست آمد که به صورت گاواژ در 5 روز در هفته در مدت 4 هفته با دوز mg/kg 100 به گروههای هدف داده شد. همچنین از تستوسترون سـنتتیک با دوز 100 میلی گرم بر میلی لیتر ساخت شرکت ایران هورمون به عنوان کنترل مثبت استفاده شد. در این مداخله رت ها پنج روز در هفته mg/kg 2 تستوسترون سـنتتیک را به صورت زیر جلدی دریافت نمودند (20).
به منظور حذف اثر حاد تمرین، 48 ساعت بعد از آخرین جلسه و پس از 12 ساعت ناشتایی، نمونه گیری خونی از گروههای پژوهش در حالت بیهوشی انجام شد. در ادامه با قربانی شدن حیوان، نخاع به دقت از بدن خارج و در محلول فرمالین 10 درصد و بافر فسفات به مدت 48 ساعت در دمای 80- درجه سانتی گراد نگهداری شد. سپس مراحل آب گیری با استفاده از الکلهای درجه بندی شده انجام و با پارافین قالب گیری شد. در نهایت برشهای 5 میکرونی ایجاد شده با دستگاه میکروتوم (مدل DS9502، شرکت دید سبز) روی لام شیشهای آغشته به چسب سیلان جهت رنگ آمیزی H&E قرار گرفت (21). میزان بیان ژنهای اوکلودین و کادهرین بافت نخاع با استفاده از کیتهای شرکت دنا- زیست آسیا طبق روش Real-time PCR ارزیابی شد. در این روش چون اسیدنوکلئیک هدف از نوع RNA بود، به یک مرحله اضافی تبدیل RNA به DNA با استفاده از آنزیم نسخه بردار معکوس 1 یا Real-time نیاز داشتیم. در مرحله آمادهسازی، ابتدا کل RNA موجود در نمونه بافت استخراج شد. سپس پرایمرهایی برای ژن اوکلودین و کادهرین از شرکت پارس توس سفارش داده شد. در مرحله بازکردن RNA با افزایش دما تا ۶۵ درجه سانتیگراد به مدت ۵ دقیقه، ساختارهای ثانویه و پیچوتابهای RNA های نمونه از همدیگر باز شد. همچنین غلظت RNA استخراج شده با استفاده از دستگاه اسپکتروفتومتری تعیین و با استفاده از آنزیم کپی برداری معکوس به cDNA تبدیل شد. در نهایت cDNA حاصل، جهت حذف مقادیر ژنومی با آنزیم DNase I تیمار و به مرحله تکثیر رسید (جدول 1). از طرفی میزان بیان پروتئینهای اوکلودین و کادهرین با استفاده از روش وسترن بلات مطابق با دستورالعمل تاتسوتا و همکاران (2015) توسط سازمان مشاوره تحقیقاتی RCO با برند تجاری هیستوژنوتک صورت گرفت. در این تکنیک برای شناسایی یک پروتئین از مخلوط پیچیدهای از پروتئینهای استخراج شده از سلول و بافت استفاده میشود. سه مرحله جداسازی براساس اندازه پروتئینها توسط ژل SDS-PAGE، انتقال به غشای کاغذی و نشاندار کردن پروتئینهای هدف با استفاده از آنتیبادی های اولیه و ثانویه فرآیند کلی این روش محسوب میشوند (22).
جدول 1. توالی پرایمرهای مورد استفاده جهت سنجش بیان ژنهای اوکلودین و کادهرین
| توالی پرایمر/ ژن |
TM |
| اوکلودین |
|
| Forward: 5´GGTGATCGGTCCCAACAAGGA -3´ |
57.1 °c |
| Reverse: 5´-CACGCTGGCACAGCCACTC -3´ |
56.4 °c |
| کادهرین |
|
| Forward: 5´-CACGGCCTTCCCTACTTCAC- 3´ |
63.5 °c |
| Reverse: 5´-TGCAAGTGCATCATCGTTGT- 3´ |
62.2 °c |
به منظور توصیف دادههای جمع آوری شده از شاخصهای گرایش مرکزی و پراکندگی و جهت بررسی طبیعی بودن توزیع دادهها از آزمون شاپیرو- ویلک استفاده شد. جهت آزمون اثر اصلی تمرین، تستوسترون سـنتتیک/ عصاره گرده خرما و اثر تعاملی تمرین و دارو بر متغیرهای وابسته از تحلیل واریانس دو طرفه استفاده گردید. در صورت مشاهده تفاوت معنادار، محل تفاوت توسط آزمون تعقیبی بونفرونی مشخص شده است. کلیه عملیات آماری با استفاده از نرمافزار SPSS نسخۀ 23 انجام و نتایج در سطح معناداری 05/0 همراه با اشکال و گرافهای حاصل از نرم افزار اکسل گزارش شده است.
یافتهها
نتایج حاصل از رنگ آمیزی بافتی به روش H&E بیانگر تغییرات بارز در بافت نخاعی گروههای تستوسترون و تعاملی شامل افزایش لیپولیز، کاهش تعداد و اندازه آدیپوست ها بود. همچنین تعداد سلولهای التهابی سیستم عصبی این گروهها بصورت معناداری کاهش داشت (05/0>P) (شکل 1). چهار هفته تمرین مقاومتی (373/0, ƞ=010/0, P=08/10=F)، دریافت تستوسترون و گرده خرما (430/0, ƞ=005/0, P=00/13=F) موجب افزایش معنادار بیان ژن اوکلودین شد. با اینحال میزان افزایش بیان این ژن درگروه DPP به طور قابل توجهی بیشتر از گروه تستوسترون سـنتتیک بود (001/0P=). همچنین تعامل تمرین مقاومتی و دارو (DPP و تستوسترون) اثر سینرژیکی بر میزان بیان این ژن داشت (672/0, ƞ=001/0, P=99/13=F). با وجودی که بیشترین میزان بیان ژن اوکلودین هنگام استفاده ترکیبی از تستوسترون سـنتتیک قابل مشاهده بود، اما از نظر آماری بین میزان اثرگذاری آن با ترکیب عصاره DPP تفاوت معناداری وجود نداشت (059/0P=) (شکل 2).
در رابطه با بیان ژن کادهرین نخاعی، تمرین مقاومتی (265/0, ƞ=015/0, P=40/6=F)، دریافت تستوسترون و گرده خرما (441/0, ƞ=004/0, P=70/9=F) سبب افزایش معنادار بیان این ژن شد. با اینحال بین میزان اثرگذاری این سه مداخله تفاوت معناداری دیده نشد (866/0P=). همچنین تعامل تمرین مقاومتی و دارو (DPP وتستوسترون) دارای اثر سینرژیکی بر بیان ژن این پروتئین بود (728/0, ƞ=001/0, P=08/15=F). بیشترین میزان بیان ژن کادهرین در هنگام ترکیب همزمان تمرین مقاومتی و عصاره گرده خرما قابل مشاهده بود. با اینحال از نظر آماری بین اثر هم افزایی عصاره DPP و تستوسترون تفاوت معناداری وجود نداشت (066/0P=) (شکل 3).
در نهایت، تمرین مقاومتی سبب تغییر معنادار میزان سنتز پروتئین اوکلودین (096/0, ƞ=404/0, P=951/0=F) و کادهرین (079/0, ƞ=647/0, P=783/0=F) نشد. با اینحال دریافت داروها (261/0, ƞ=021/0, P=29/5=F); (252/0, ƞ=033/0, P=08/5=F) و تعامل تمرین مقاومتی با داروها (839/0, ƞ=001/0, P=318/8=F); (862/0, ƞ=001/0, P=624/8=F) بیان این پروتئینها را به ترتیب افزایشی کرد. نتایج آزمون بن فرونی نشان داد اثرگذاری مداخله دارو تنها مربوط به تستوسترون سـنتتیک بوده است. از این رو، بین میزان اثرگذاری گرده خرما و تستوسترون بر بیان پروتئین اوکلودین (016/0P=) و کادهرین (012/0P=) نخاعی تفاوت معناداری وجود داشت. در هر دو پروتئین علی رغم قابل مشاهده بودن اثر سینرژیک تمرین مقاومتی و مکمل DPP در بیان پروتئینها؛ بیشترین میزان سنتز اوکلودین (026/0P=) و کادهرین (017/0P=) هنگام تعامل تمرین مقاومتی و مکمل تستوسترون مشاهده شد (شکل 4).
 شکل 1. فتومیکروگراف نوری بافت نخاع گروهها با رنگ آمیزی هماتوکسیلین- ائوزین با بزرگ نمایی ×100 (100 µm)
شکل 1. فتومیکروگراف نوری بافت نخاع گروهها با رنگ آمیزی هماتوکسیلین- ائوزین با بزرگ نمایی ×100 (100 µm)
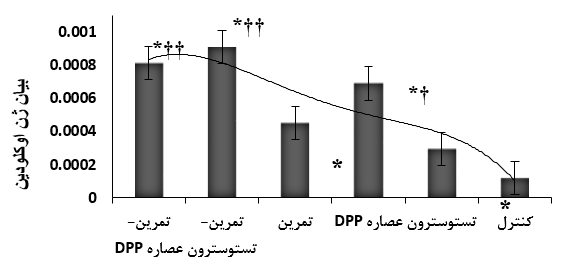 شکل 2. اثرات تمرین مقاومتی، تستوسترون و عصاره گرده خرما بر بیان ژن اوکلودین سد نخاعی- خونی رت های نر
* نشانه افزایش معنادار نسبت به گروه کنترل، †* نشانه افزایش معنادار نسبت به گروه تمرین و تستوسترون، ††* نشانه افزایش معنادار نسبت به گروه کنترل، تستوسترون، عصاره DPP و تمرین مقاومتی
شکل 2. اثرات تمرین مقاومتی، تستوسترون و عصاره گرده خرما بر بیان ژن اوکلودین سد نخاعی- خونی رت های نر
* نشانه افزایش معنادار نسبت به گروه کنترل، †* نشانه افزایش معنادار نسبت به گروه تمرین و تستوسترون، ††* نشانه افزایش معنادار نسبت به گروه کنترل، تستوسترون، عصاره DPP و تمرین مقاومتی
 شکل 3. اثرات تمرین مقاومتی، تستوسترون و عصاره گرده خرما بر بیان ژن کادهرین سد نخاعی- خونی رت های نر
* نشانه افزایش معنادار نسبت به گروه کنترل، †* نشانه افزایش معنادار نسبت به گروه تمرین، عصاره DPP و تستوسترون
شکل 3. اثرات تمرین مقاومتی، تستوسترون و عصاره گرده خرما بر بیان ژن کادهرین سد نخاعی- خونی رت های نر
* نشانه افزایش معنادار نسبت به گروه کنترل، †* نشانه افزایش معنادار نسبت به گروه تمرین، عصاره DPP و تستوسترون

شکل 4. اثر تمرین مقاومتی، تستوسترون و عصاره گرده خرما بر بیان پروتئین اوکلودین و کادهرین سد نخاعی- خونی رت های نر.
در هر دو پروتئین: * افزایش معنادار نسبت به گروه کنترل، عصاره DPP و تمرین مقاومتی †* افزایش معنادار نسبت به گروه کنترل، تستوسترون، عصاره DPP، تمرین مقاومتی و تمرین- عصاره DPP
بحث
هدف از مطالعه حاضر، بررسی اثرات مستقل و تعاملی تمرین مقاومتی، تستوسترون سـنتتیک و عصاره گرده خرما بر بیان ژن و سنتز پروتئینهای اوکلودین و کادهرین سد نخاعی- خونی موشهای صحرایی نر جوان بود. در این راستا ارزیابی بافت شناسی با استفاده از رنگ آمیزی هماتوکسیلین- ائوزین نشان داد میزان نفوذپری سد BSCB رت های گروه تستوسترون و ترکیبی نسبت به سایر گروهها کاهش و یکپارچگی اتصالات اندوتلیال آنها افزایش یافته است. احتمالاً یکی از علل این اثر، کاهش حضور سلولهای التهابی در بافت سیستم عصبی و عدم توانایی آنها در مهاجرت از سدها باشد (23). همچنین براساس نتایج ما افزایش یکپارچگی سد خونی- نخاعی میتواند ناشی از افزایش بیان پروتئینهای اتصالات محکم و اتصالات چسبنده باشد. در واقع یکپارچگی این سد وابسته به اتصالات محکمی است که پروتئینهای تخصصی آن (ZO-1، ZO-2، کلودین- 5 و اوکلودین) مسئول حفظ ساختار سه بعدی هستند (24). از طرفی اختلال در تولید و میزان بیان کادهرین به عنوان یکی از اتصالات چسبنده سد از علل مهم بروز بیماریهای متابولیکی و سرطان است. از اینرو وجود این پروتئینهای تخصصی در بافت ضروری بوده و اختلال در عملکرد ژنی آنها با ایجاد اختلال در سنتز نوع پروتئین نقش مهمّی در عفونت باکتریایی، التهاب، آپوپتوز و پیشرفت تومورها دارند (25). بر مبنای ادبیات موجود، استفاده از مکملهای گیاهی یا فعالیتهای ورزشی احتمالاً راهکارهای مؤثری در تنطیم سطوح این پروتئینها هستند. این شواهد نشان میدهد که انتخاب سبک زندگی قادر به تنظیم سد BSCB از لحاظ ساختاری و عملکردی است.
نتایج ما نشان داد چهار هفته تمرین مقاومتی، میزان بیان ژن پروتین اوکلودین سد خونی- نخاعی را نسبت به گروه کنترل افزایش داده است. این پژوهش اوّلین مطالعهای است که افزایش بیان ژن پروتئینهای تخصصی سد نخاعی آزمودنیهای سالم متعاقب تمرین مقاومتی نشان میدهد؛ زیرا قبلاً تنها اثر این تمرینات در بیماران عصبی ارزیابی شده است. الجنگی و همکاران در یک بررسی علمی نشان دادند تمرینات قدرتی و استقامتی علاوه بر حفظ پروتئینهای TJ بافت نخاع، از طریق محدود کردن ورود سلولهای خودواکنشی T به CNS، یکپارچگی BBB بیماران MS را ارتقا میدهند (26). اینکه فعالیتهای هوازی از طریق تنظیم آنزیمهای فسفوریلاسیون پروتئینهای کلودین-1 و اوکلودین نقش مهمّی در جلوگیری از آپوپتوز سلولهای سالم روده یا افزایش آپوپتوز سلولهای سرطانی دارند، کاملاً مشخص است (27). از طرفی فعالیتهای قدرتی احتمالاً از طریق بهبود سطح تماس عروق مغزی و افزایش عملکرد متابولیک سلولهای اندوتلیال، میزان استرس اکسیداتیو نورونهای عصبی را کاهش میدهند. به علاوه این تمرینات با افزایش سطح فاکتور نوروتروفیک مشتق از مغز (BDNF) و فاکتور رشد شبه انسولین منجر به افزایش غلظت سایتوکین های ضد التهابی و کاهش سایتوکین های پیش التهابی میشوند (28). این نتایج در راستای تحقیقاتی هستند که نشان دادهاند استرس اکسیداتیو با تغییر ساختار و موقعیت کلودین ها انسجام سدها را مختل میکنند. همچنین براساس نتایج جمع بندی شده مالکیویچ و همکاران، فعالیت بدنی از طریق کنترل عواملی از جمله سیستم رنین- آنژیوتانسین و نورآدرنرژیک مغز، عملکرد اتونوم مرکزی و مسیر کینورنیناین سد BBB را تحت تأثیر قرار میدهد (29). این نتایج نشان دهنده نقش بالقوه شرطی سازی تمرینات ورزشی بر مؤلفههای اتصالات محکم است که در نهایت منجر به بهبود عملکرد اندوتلیال و افزایش تراکم مویرگهای سیستم عصبی میشود. اینکه کدامیک از سایتوکین های ضد التهابی با تنظیم بیان اوکلودین میزان اختلال اتصالات محکم مغز را کاهش و عملکرد سد خونی- نخاعی را بهبود میبخشند، باید در مطالعات آینده بررسی شود. از سوی برخی محققان معتقدند فقط ورزش اجباری از جهت امکان دستکاری شدت، مدت و تعداد تکرار، پروتکل مفیدی در محافظت عصبی و تغییر متابولیسم مغز است (16, 21). با اینحال یعقوبی و همکاران نشان دادند هر دو تمرین ورزش اجباری و اختیاری ابزار مفیدی برای CNS از جهت افزایش بیان ژن دو پروتئین اوکلودین و کلودین 5 هستند (30).
جدیدترین یافته ما نشان داد تمرین مقاومتی علاوه بر افزایش بیان ژن پروتئینهای اتصالات محکم سد نخاعی- خونی باعث افزایش بیان ژن مولکول چسبنده یا اتصالی کادهرین میشود. انتظار میرود ضمن این اتفاق سطوح RNA طیفی از پروتئینهای دیگر سد افزایش یافته باشد که طبیعتاً سبب افزایش ناقلین برون ریز ژن از جمله MDR1 خواهد شد (31). علیرغم این نتایج دلگرم کننده، چندین مطالعه اثرات منفی ورزش طولانی مدت بر میزان نفوذپذیری BBB را گزارش کردهاند. از نظر این پژوهشگران فعالیت بدنی شدید و طولانی مدت با افزایش آنزیمهای کلیدی فسفوریلاسیون، عملکرد پروتئینهای اتصالی را با اختلال مواجه میکند (9). اگرچه هنوز الگو و معیار مناسبی برای مقایسه این یافتهها وجود ندارد، احتمالاً علت تفاوت در نتایج مربوط به تفسیر نادرست تغییرات بیان ژن و پروتئین و ارتباط آن با میزان نفوذپذیری باشد. در این زمینه برخی از پژوهشگران افزایش بیان ژن را معادل کاهش نفوذپذیری تفسیر کردهاند (32)؛ در حالیکه در این زمینه باید افزایش سنتز پروتئین را معیار قرار داد. نتایج ما نشان میدهد 4 هفته تمرین مقاومتی منحصراً میتواند منجر به افزایش بیان ژن پروتئینهای اوکلودین و کادهرین شود. در واقع اثرگذاری این مداخله به مرحله بیان پروتئین یا مرحله بالفعل کاهش نفوذپذیری سد نرسیده است. از طرفی خدادوست و همکاران به این نتیجه رسیدند که تمرینات ورزشی تنها بر نشانگرهای نفوذپذیری سد خونی- مغزی و وضعیت فاکتور نروتروفیک آزمودنیهای نر دارای وزن طبیعی تأثیرگذار است (33). همچنین شواهد روز افزونی مبنی بر افزایش قابلیت نفوذ سد خونی- نخاعی در شرایط متغیر توأم با افزایش گرما وجود دارد (34). این دادهها حاکی از این است افزایش گرما مستقل از علت آن اثرات قابل توجهی بر سلولهای عصبی، گلیال، اندوتلیال و کارکرد سدها دارد. هرچند در این پژوهش دمای بدن رت های گروه تمرینی اندازه گیری نشده است؛ میتوان فرض کرد شدت 4 هفته تمرین مقاومتی به اندازهای بوده است که باعث افزایش دما و کنترل بیان پروتئینهای اوکلودین و کادهرین شده است.
در خصوص مکمل تستوسترون سـنتتیک یافتههای این مطالعه نشان داد 4 هفته مصرف آن موجب تغییرات افزایشی میزان بیان ژن اوکلودین و کادهرین در مقایسه با گروه کنترل شده است. با اینحال، این مکمل برخلاف گرده خرما و تمرین مقاومتی قادر به افزایش معنادار نمود پروتئینهای اتصال محکم و چسبنده نخاعی نیز شده است. این یافته نشان دهنده این است که مکمل تستوسترون میتواند جایگزین مناسبی برای افزایش سنتز پروتئینهای سد خونی- نخاعی و کاهش نفوذپذیری آن در آزمودنیهای سالم باشد. تا امروز پژوهشهای محدودی اثرات هورمونهای استروئیدی را بر عملکرد عروق مغزی بررسی کردهاند. براساس دادههای موجود تستوسترون قادر به تحریک بیان پروتئینهای کلودین،ZO-1 و JAM-A سد خونی- مغزی و سد بیضه- خون (BTB) موشهای نر از طریق مکانیسمهای ژنی و غیرژنی سلولهای اندوتلیال عروقی است (35). از طرفی طی دوران یائسگی یا در آستانه آن کاهش انسجام سد خونی- مغزی با تغییر هورمونهای جنسی پلاسمای خون ارتباط معناداری نشان میدهد (36). با اینکه از نظر محققان تأثیر نهایی تستوسترون بر عروق عصبی احتمالاً ناشی از قابلیت متابولیک بیشتر دهیدروتستوسترون (DHT) است؛ ظاهراً اثر تستوترون سـنتتیک بر اندوتلیال های خونی- نخاعی وابسته به افزایش هورمونهای در گردش یا ایجاد پیوند موضعی است (37). رحمتی و همکاران در بررسی اثر مداخلات نشان دادند کاهــش نفوذپذیــری ســد خونی- مغــزی ناشی از کاهش بیـان مقادیـر مولکولهای چسـبان (ICAM-1 و VCAM-1) است (38). از سویی در سد خونی- نخاعی پروتئین P-gp یک پمپ انتشار مهم برای تعدادی از هورمونهای استروئیدی است. در این زمینه برخی محققان معتقدند هورمونهای پروژسترون و استرادیول باعث افزایش ظهور P-gp از طریق قاعده نسخه برداری میشوند. بر این اساس امکان دارد تنظیم بالای این پروتئین علامتی برای کاهش قابلیت نفوذپذیری سد BSCB موشهای نر سالم باشد (35, 37). با اینحال در این پژوهش میزان تنظیم پروتئین P-gp سد نخاعی- خونی گروههای آزمایشی مورد بررسی قرار نگرفته است.
یافتههای قبلی نشان میدهد تزریق دوزهای بیش از حد تستوسترون میزان نمود پروتئینهای اندوتلیال سیستم اعصاب مرکزی و به همین نسبت عملکرد سد خونی- مغزی را تحت تاثیرگذار قرار میدهد (35). بنابراین اثر این مکمل احتمالاً وابسته به دوز مصرفی است. در این رابطه، نیروینسکا و همکاران با بررسی اثر تمرین استقامتی (6 هفته به مدت 60-40 دقیقه، 5 بار در هر هفته) و مکمل تستوسترون (80 میلی گرم/ کیلوگرم وزن بدن) گزارش دادند تستوسترون سطوح کلودین- 5، اوکلودین و کادهرین سد خونی- نخاعی را صرف نظر از تمرین استقامتی کاهش میدهد (32). بنابراین تغییرات واقعی پروتئینهای تحلیل شده سد نخاعی- خونی پیچیدهتر از آنچه چیزی است که قبلاً تصور میشد. تا آنجا که می دانیم، هنوز دلایل معتبری برای رفع تناقضهای مربوط به بینشهای مولکولی و بالینی وجود ندارد. در این راستا برای نتیجه گیری بهتر و جهت گیری مطالعات آینده یک بررسی فراتحلیلی از حوزههای مختلف تحقیقات پزشکی مورد نیاز است. اینکه عامل تعیین کننده در این زمینه نوع بافت مورد بررسی است یا خیر و تداخل ایجاد شده در مسیرهای سنتز پروتئینهای اوکلودین و کادهرین ناشی از نوع مداخله بوده است، باید در کارآزمایی های بالینی دقیق مورد بررسی قرار گیرد.
دیگر یافته مطالعه حاضر نشان داد عصاره گرده خرما اثرگذاری بیشتری نسبت به تمرین مقاومتی و تستوسترون در تقویت بیان ژن پروتین های سد نخاعی- خونی موشهای نر دارد. این اثر افزایشی در رابطه با بیان ژن اوکلودین معنادار و در رابطه با بیان ژن کادهرین غیرمعنادار بوده است. بنابراین فرضیه استفاده از گرده نـر نخـل خرمـا بـه عنوان یک ترکیب طبیعی جایگزین تقویـت میشود (39). از سویی این عصاره اثر معناداری بر بیان پروتئینهای اوکلودین و کادهرین نداشت. از آنجایکه مطالعات مشابهی در این زمینه یافت نشد، تببین نتایج ما براساس مکانیسمهای دقیق سلولی و مسیرهای بیان ژن دشوار است. با اینحال، عابدی و همکاران با بررسی مطالعات مروری مربوط به عصاره گرده خرما گزارش کردند این عصاره با دارا بودن انواع اسید آمینهها و ویتامینهای مفید، اثرات مثبتی بر هورمونهای جنسی نمونههای انسانی و حیوانی دارد (40). از طرفی نشان داده شده است رادیکالهای آزاد باعث اختلال در عملکرد اتصالات محکم میشوند. بنابراین، تقویت سیستم آنتی اکسیدانی بدن یکی از مکانیسمهای توجیه کننده اثر این مداخله بر بیان ژن پروتئینهای سد است. احتمالاً اثرات مفید این گرده بروی مارکرهای التهابی و استرس اکسیداتیو به درصد بالای آلکالوئیدها، تانن ها، فلاونوئیدها، تـرپن هـا و قنـدها مربوط است. همچنین ترکیباتی نظیر روی و کادمیوم میتوانند از طریق افزایش بیوسنتز 17- بتا هیدورکسی دهیدروژناز باعث افزایش متابولیسم استروئیدها و پروتئینها گردند (41). اینکه کدام یک از ترکیبات گرده خرما مسئول تنظیم بیان ژنی پروتئینهای سد نخاعی- خونی موشهای نر است، باید در مطالعات بعدی مشخص شوند. از نظر بعضی محققان بیان پروتئین و ژنهای مرتبط با آپوپتوز در پاسخ به مصرف گرده نخل کاهش پیدا خواهد کرد (41, 42). بنابراین افزایش بیان ژنی پروتئینها متعاقب استفاده از عصاره DPP شاید مکانیسمی عمل کننده علیه آپوپتوز سلولهای اندوتلیال سد نخاعی- خونی باشد. در این راستا اندازه گیری سیتوکین های التهابی و شاخصهای آپوپتوز همراه با تعیین ارتباط آنها با پروتئینهای اتصال محکم و چسبنده به یافتن مکانیسمهای اثرگذار گرده نخل خرما کمک شایانی خواهد کرد.
در نهایت نتایج حاصل از این مطالعه بیانگر اثرات سینرژیک تمرین مقاومتی/ تستوسترون و عصاره گرده خرما/ تمرین مقاومتی در افزایش بیان ژن و بیان پروتئین اوکلودین و کادهرین سد نخاعی خونی بود. نکته جالب توجه عدم وجود تفاوت معنادار بین میزان تأثیرگذاری ترکیب این دو مداخله بر بیان ژنها و برتری ترکیب تمرین مقاومتی/ تستوسترون در بیان یا سنتز پروتئینها بود. هم راستا با این نتایج، تحقیقات همسویی در رابطه با اثرات اثر هم افزای ترکیب عصاره گرده خرما با تمرین قدرتی بر سطوح گیرندههای هورمون لوتئینی و بیان ژن سایر پروئین های اتصال محکم مانند کلودین 1 وجود دارد (13, 19). به نظر میرسد مکانیسم عمل این مداخله بواسطه تعدیل اختلال در نشانگرهای حیاتی یا ارتقای سطوح پروتئینهای کارکردی باشد. از طرفی بیشترین افزایش بیان پروتئین اوکلودین و کادهرین و متعاقباً آن بیشترین کاهش نفوذپذیری سد نخاعی- خونی هنگام ادغام تمرین مقاومتی با مکمل تستوسترون، تنها براساس تأثیر قابل توجه تستوسترون قابل توجیه است. در این زمینه نیروینسکا و همکاران نشان دادند تمرین استقامتی در مقایسه با دو دوز تستوسترون اثر معناداری بر سطح پروتئینهای اندوتلیال بافت نخاعی موشها ندارد (32). همچنین نظریان و همکاران با بررسی اثر عصاره گرده نخل (100 میلی گرم به ازای هر کیلوگرم وزن بدن)، تستوسترون سـنتتیک (2 میلی گرم به ازای هر کیلوگرم وزن بدن) و تمرین مقاومتی (5 روز در هفته به مدت چهار هفته) بر پروتئینهای عامل اتصالات محکم بافت پروستات موشهای صحرایی نر نشان دادند استفاده همزمان از تمرین و تستوسترون/ گرده نخل اثر هم افزایی بر بیان پروتئین اوکلودین دارد (20). اربطی و همکاران (2021) مکانیسم اثرگذاری این مداخلات را در بافت چربی قهوهای موشهای صحرایی نر مربوط به افزایش بیان ژنهای آدیپونکتین، افزایش فعالیت SOD و کاهش فعالیت MDA دانستهاند. در این شرایط احتمالاً سطوح فعالیت GPx و GSH نیز کاهشی میشود (19). اگرچه این نتایج برای بیماریهای مرتبط با نفوذپذیری سد نخاعی- خونی دارای اهمیت کاربردی است، بررسی سایر پروتئینهای مجموعه سد در پژوهشهای آینده ضروری است. مرور تحقیقات نشان میدهد میزان تغییر بیان ژن و سنتز پروتئین سدهای عصبی عموماً متفاوت از یکدیگر است. از طرفی بررسی نمود پروتئینهای سد نخاعی- خونی با روش وسترن بلات تنها موقعیت نسبی آنها را در یک ترکیب همگن مشخص میکند. از این رو، باید میزان تغییر پروتئینهای غیرسدی بافت نیز در ارزیابیهای سلولی مد نظر قرار گیرد. با توجه به اینکه BSCB معادل عملکردی BBB است، جهت تفسیر منطقی و تأیید این نتایج به پژوهشهایی درباره نقـش کمکـی تمرینات ورزشی و دوزهای عصاره DPP در کاهش نفوذپذیری دو سد نیاز داریم.
نتیجه گیری
یافتههای پژوهش حاضر میزان تغییر فیزیولوژیکی قابل برگشت بیان ژن و بیان پروتین های اوکلودین و کادهرین سد BSCB را پس از چهار هفته تمرین مقاومتی و مصرف عصاره DPP/ تستوسترون نشان داد. ظاهراً این مداخلات اثرگذاری بیشتری بر بیان ژن پروتئینهای اتصال محکم نسبت به اتصالات چسبنده دارند. با اینحال روند اثرگذاری آنها بر بیان پروتئینهای اوکلودین و کادهرین کاملاً مشابه است. علی رغم اینکه عصاره DPP میتواند جایگزین مناسبی برای افزایش بیان ژن پروتئینها نسبت به تستوسترون و تمرین مقاومتی باشد، مصرف مجزای تستوسترون از لحاظ افزایش بیان پروتئینهای اوکلودین و کادهرین دارای برتری است. بنابراین استفاده منفرد از تستوسترون با دوز 2 میلی گرم/ کیلوگرم وزن بدن باعث کاهش نفوذپذیری سد نخاعی- خونی نمونههای نر سالم حیوانی میشود. با اینحال، جهت افزایش اثر سینرژیک، ترکیت تمرین مقاومتی با تستوسترون اولویت بیشتری دارد.
تشکر و قدردانی
این مقاله بخشی از نتایج رساله دکتری با شماره طرح 148000602913700017162703571 و کد اخلاق IR.UT.SPORT.REC.1397.028 در سال 1401 بوده که به تأیید معاونت پژوهشی دانشگاه آزاد واحد تهران مرکزی رسیده است. مراتب قدردانی خود را از همکاران و اساتید محترم اعلام میداریم.
نوع مطالعه:
پژوهشي |
موضوع مقاله:
فیزیولوژی








